- C end of Torus and N end of RRM linker
- Loop (residues 116-133) with a6 at its end

Figure
1. Structure of connector loop in Cwc2, conserved positively charged residues
highlighted
- Involved in RNA recognition? ~ Conserved residues (compare to RNA motifs/complementarity to RNA)
The possibility of the
connector loop being involved in RNA recognition was explored by carrying out a
BLAST on its sequence.

Figure 2. BLAST
results for connector loop
We can see that we are
not getting results with known proteins with RNA-binding motifs. Most of the
results are hypothetical proteins characterised from open reading frames.
Perhaps further research on possible domains of these proteins might lend
insight into the possibility of the connector loop interacting RNA. Evidence
for this was obtained from mutagenesis experiments on the connector loop. (Lu
et al, 2012)
However, the loop
might still function to guide RNA to the depression (pocket) and this is explored
below.
- Contains positively charged patch, interconnected to patches on RRM and ZnF

Figure 3. Electrostatic surface potential of Cwc2
The blue patches show
the positively charged regions on the connector loop, around the ZnF and the
RRM (from top to bottom). The patches are linked by positively charged residues.
The patches line depression and could be said to hold RNA secondary structure. This
charge distribution on the surface could be vital to guiding the negatively
charged RNA in the right conformation to the binding sites, thereby making
binding more efficient.
- Connector form rear rim and is on top of depression ~ guide RNA to pocket?

We see that there are
many potential RNA binding sites predicted and are present in the connector
loop as well. However this is only a rough gauge and further experimentation
involving isolated connector loop and RNA binding assays needs to be carried
out. But we can say with some confidence the connector loop is involved in
guiding the RNA to the binding pocket, thereby increasing efficiency of Cwc2.
- Crosslinks with U6 snRNA ~ all contacts: ZnF (C87), the connector element (Y120–R121), the RRM (K152, C181 from RNP1 and Y138 from RNP2) and in the Torus domain (F47)

Figure 4.
Binding site to U6 snRNA characterised by crosslinking highlighted residues
We can see from above
that the shown top view face of the molecule is implicated in binding and assisting
in splicing to form mature U6 snRNA. Other literature indicates the importance
of Cwc2 in this function. Furthermore it is seen that U6 snRNA is not the only
snRNA involved in regulation by Cwc2.

Figure 5. Gel run of
glucose induced Cwc2 gene-repressed yeast cell extract (Mcgrail. et al, 2009
Upon switching from
inducing to repressing Cwc2 expression under Gal1 promoter, we see that the
levels of U1, U4, U5 and U6 snRNA levels drop indicating possible interactions
in these snRNAs with Cwc2.
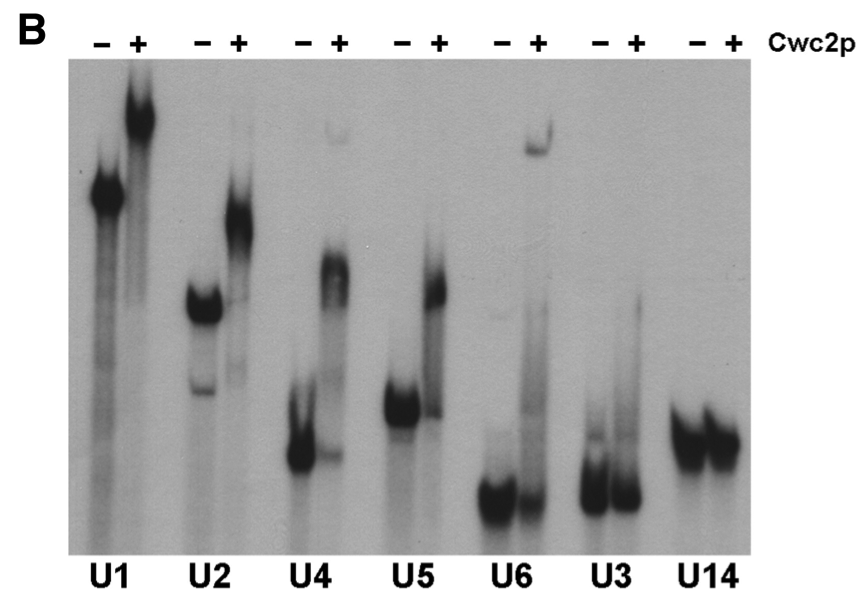
Figure 6. Gel shift
assay using non-denaturing PAGE with snRNA and Cwc2
We see that there is specific binding and slower movement of complex by
Cwc2 binding to U1, U2, U4 and U5 snRNA. The results shown in Figure 5 are
supported by Figure 6 results. Furthermore, Cwc2 was shown
to crosslink with U6 snRNA using immunoblotting with anti-TAP antibody against
TAP-tagged (tandem affinity purification) Cwc2. Cwc2-TAP crosslinked to U6
snRNA in the first and second steps of spliceosome formation in yeast. Thus we
can postulate that Cwc2 functions in binding U6 to spliceosome upon U4
dissociating and directing its interactions with pre-mRNA. (McGrail et al,
2009) It
was also found that the connector and ZnF bind U6 than U4 snRNA with greater
affinity. The whole complex serves to carry out pre-mRNA splicing activity and
Cwc2 plays an important role in mediating it. This is seen by the point
mutation Y120A removing splicing activity. We can thus tell that the
hydrophobic interactions set up by the tyrosine-120 residue is key to
positioning spliceosome components for its activity.
- Connector loop is similar to nuclear cap-binding complex CBC20 N terminal extension (Mazzaet al, 2002)

Figure 7. Alignment of
CBC20 with Cwc2 (CBC in orange and Cwc2 in blue)
There is conservation
that is reflected in the amino acid sequence to an extent as well. We can see
that the alpha helices are roughly in the similar positions above. This gives
further proof that the novel structure of Cwc2 is evolutionarily conserved and
therefore advantageous in carrying out its function of RNA binding. The
important Y120 residue on cwc2 connector loop is conserved in CBC at
tyrosine-20 position on its N-terminal extension. Y20 on CBC could not be
visualised above as the CBC80 subunit structure was used. Further work on CBC
will allow us greater insight into mechanism of binding.

Figure 8. Structure of
CBC20 subunit with key residues that interact cap highlighted
The yellow residues in
Figure 8 are shown in cap-bound conformation whilst the blue residues undergo
conformational change to accommodate the cap. The N-terminal extension allows
high flexibility much like the connector loop and is key to the functioning of CBC.
- high flexibility, many conformational states ~ important to function as molecular switch (molecular dynamics simulation CHARMM)
The high flexibility
of the connector loop can be further characterised in terms of its molecular
dynamics. A simulation program such as CHARMM can be run to look at the extent
of flexibility the connector loop can accommodate.
- Molecular switch (accommodate shifting contacts in spliceosomal transitions)

Figure 9. Ramachandran plot of whole Cwc2
protein (drawn using RAMPAGE)

Figure 10. Ramachandran plot for Connector loop (residues 116-133) using Ramachandran Plot 2.0 program
We see that generally there is much allowed
flexibility in terms of torsions and phi/psi angles for the whole molecule and
for the connector loop specifically too. However this is a very cursory view
and further analysis should be carried out by drawing plots using MATLAB based
on PDB data. However we can tell
that the conformations of the loop does not change so quickly that the electron
density cloud could not be fixed enough for x-ray crystallography
visualization. We can conclude that there are different conformational states
associated with the connector loop and that this property might be integral to
Cwc2 function.
No comments:
Post a Comment